
| V: Bioelektrizität (INHALT) |
|
| LOWRES 35 kbyte |
| Das Riesenaxon des Tintenfischs |
|---|
 Für die Erforschung physiologischer Phänomene wie der Bioelektrizität gibt es einige
wichtige Präparationen. Die grundlegenden Informationen über die Nervenleitung sind am
Riesenaxon des Tintenfischs Loligo peali gewonnen worden. Diese Tiere brauchen zum Überleben
einen sehr schnellen Fluchtreflex, der auf einer blitzschnellen Muskelkontraktion beruht. Der wie ein
Muskelschlauch ausgebildete, nur am Kopfenende offene Mantel wird dazu mit Wasser gefüllt. Die Kontraktion
des Mantels wird dann von kopfnahen Ganglien ausgelöst, und erfolgt so, daß der Mantel in seiner
ganzen Länge gleichzeitig kontrahiert. Dazu ist es notwendig, daß das Nervensignal sehr schnell
die hinteren Teile des Mantels erreicht.
Für die Erforschung physiologischer Phänomene wie der Bioelektrizität gibt es einige
wichtige Präparationen. Die grundlegenden Informationen über die Nervenleitung sind am
Riesenaxon des Tintenfischs Loligo peali gewonnen worden. Diese Tiere brauchen zum Überleben
einen sehr schnellen Fluchtreflex, der auf einer blitzschnellen Muskelkontraktion beruht. Der wie ein
Muskelschlauch ausgebildete, nur am Kopfenende offene Mantel wird dazu mit Wasser gefüllt. Die Kontraktion
des Mantels wird dann von kopfnahen Ganglien ausgelöst, und erfolgt so, daß der Mantel in seiner
ganzen Länge gleichzeitig kontrahiert. Dazu ist es notwendig, daß das Nervensignal sehr schnell
die hinteren Teile des Mantels erreicht.
Die Geschwindigkeit der Signalleitung in einfachen (nicht-myelinisierten) Axonen hängt vom Axondurchmesser ab: je größer der Durchmesser desto schneller die Signalleitung. Beim Tintenfisch sind aus diesem Grund die Axone, die den hinteren Teil des Mantels versorgen, besonders dick. Tatsächlich gehören die Nervenzellen im Mantel von Loligo zu den größten Neuronen überhaupt, mit Axonen, die bis 10 cm lang und 1 mm dick sind. Sie lassen sich experimentell gut handhaben, perfundieren und mit Elektroden untersuchen. Mehr zu Loligo peali |
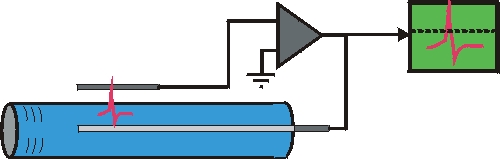 Bei den bahnbrechenden Experimenten von Hodkin und Huxley wurden Abschnitte dicker Axone vom Tintenfisch
freipräpariert, das Axoplasma wurde entfernt, und das Axon innen mit einer Salzlösung perfundiert.
Mit den Ableitungskonfigurationen der Spannungsklemme und Stromklemme (rechts ist
eine einfache Form der Sromklemme gezeigt) wurde die ionische Zusammensetzung der Aktionsströme
aufgeklärt.
Bei den bahnbrechenden Experimenten von Hodkin und Huxley wurden Abschnitte dicker Axone vom Tintenfisch
freipräpariert, das Axoplasma wurde entfernt, und das Axon innen mit einer Salzlösung perfundiert.
Mit den Ableitungskonfigurationen der Spannungsklemme und Stromklemme (rechts ist
eine einfache Form der Sromklemme gezeigt) wurde die ionische Zusammensetzung der Aktionsströme
aufgeklärt.
|
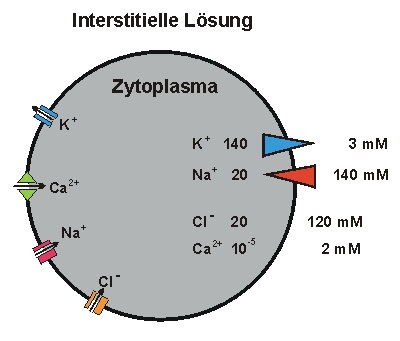 Die Grundlage aller Bioelektrizität ist die Ungleichverteilung von Ionen über der Plasmamembran
sowie die Aktivität von Ionenkanälen, die spezifisch für jeweils eine Ionensorte sind. Insbesondere
die Verteilung von Na und K sind entscheidend: Ein einwärts gerichteter Na-Gradient (rot) treibt den
depolarisierenden Na-Strom in der ersten Phase des Aktionspotentials. Ein auswärts gerichteter
K-Gradient (blau) treibt den hyperpolarisierenden K-Strom, der das Aktionspotential beendet.
Die Grundlage aller Bioelektrizität ist die Ungleichverteilung von Ionen über der Plasmamembran
sowie die Aktivität von Ionenkanälen, die spezifisch für jeweils eine Ionensorte sind. Insbesondere
die Verteilung von Na und K sind entscheidend: Ein einwärts gerichteter Na-Gradient (rot) treibt den
depolarisierenden Na-Strom in der ersten Phase des Aktionspotentials. Ein auswärts gerichteter
K-Gradient (blau) treibt den hyperpolarisierenden K-Strom, der das Aktionspotential beendet.
Sowohl die Rolle der einzelnen Ionensorten beim Zustandekommen des Aktionspotentials als auch die die Aktivität von Ionenkanälen, die die Membranspannung beeinflussen (selbst aber auch von der Membranspannung kontrolliert werden, wurden durch Studien am Riesenaxon des Tintenfischs aufgeklärt. Vorlesungskript: Ionenkanäle |