Elektronenmikrographie von:
Walter H. Schröder
Forschungszentrum Jülich
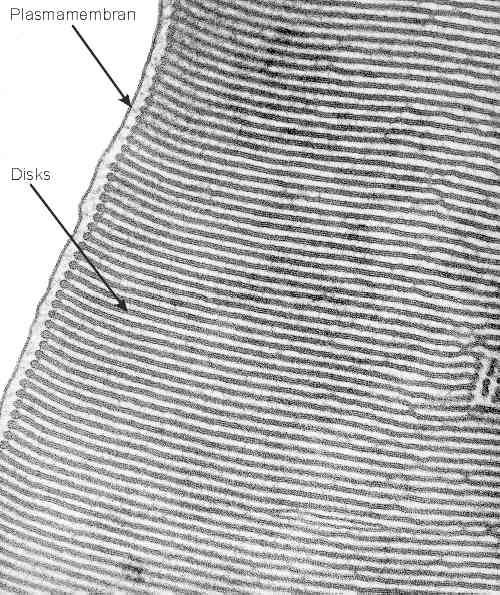
| Die Membranen des Außensegments | HIGHRES (121 kbyte) |
|---|---|
| Elektronenmikroskopische Aufnahme eines Längsschnitts durch
ein Stäbchenaußensegment. Erkennbar ist die Plasmamembran und die davon
isolierte Membran der Disks. Der Abstand zwischen Plasmamembran und Disks entspricht etwa 50 nm.
Das lichtempfindliche Pigment Rhodopsin ist in
die Membranen der Disks eingebaut und zwar in einer extrem hohen Dichte
(ca 30000 pro mm2). Wegen dieser hohen Packungsdichte
wird ein Photon, das sich entlang der Längsachse des Außensegments bewegt
(auf der Abbildung von unten links nach oben rechts) mit hoher Wahrscheinlichkeit
auf ein Rhodopsinmolekül treffen. Das ist eine wichtige Voraussetzung für die
Fähigkeit des Auges, einzelne Photonen wahrzunehmen. Elektronenmikrographie von: Walter H. Schröder Forschungszentrum Jülich |
|
| Die photo-chemische Transduktion | HIGHRES (109 kbyte) |
|---|---|
| Bei Absorption eines Photons durch Rhodopsin wird das Lichtsignal zunächst
in ein chemisches Signal umgewandelt. Das Photon wird durch Retinal absorbiert, der
Chromophor des Rhodopsinmoleküls. Eine lichtinduzierte Isomerisierung des Retinals
aktiviert Rhodopsin (Rho*).Rho* seinerseits aktiviert das GTP-bindende
Protein Transducin, das aus drei Untereinheiten (a,
b und g) besteht. Bei Aktivierung
dissoziiert die GTP-bindende a-Untereinheit von dem
bg-Komplex und aktiviert das Enzym Phosphodiesterase (PDE).
PDE hydrolysiert cGMP, den zellulären Botenstoff ("second messenger") der Phototransduktion.
Die erste Folge der Belichtung eines Photorezeptors ist also ein Abfall der cGMP-Konzentration
im Außensegment. Diese biochemische Transduktionskaskade ist sehr effizient. Die Aktivierung von einem Rhodopsinmolekül kann zur Aktivierung von 3000 PDE-Molekülen führen. Jedes davon kann bis zu 2000 cGMP-Moleküle spalten. Daraus ergibt sich theoretischer Verstärkungsgrad von 6 Millionen cGMP pro Photon. |

|